 Migrating waders in Roebuck Bay, Western Australia http://en.wikipedia.org/wiki/Bird_migration Food is the primary motivation, for birds migration; some hummingbirds will not migrate if fed through the winter. As the days shorten in autumn, the birds return to warmer regions and enjoy the constant food supply. The primary physiological cue for migration, are the changes in the day length. These changes are also related to hormonal changes in the birds. In the period before migration, many birds display higher activity as well as physiological changes such as increased fat deposition. The routes taken on forward and return migration are often different. A common pattern in North America is clockwise migration. The specific routes may be genetically programmed or learned to varying degrees. Species that move short distances, however, may not need such a timing mechanism, and may move in response to local weather conditions. Many, if not most, birds migrate in flocks. For larger birds, flying in flocks reduces the energy cost. Geese in a V-formation may conserve 12–20% of the energy they would need to fly alone. However, radar studies showed some birds; Red Knots and Dunlins, are able to fly 5 km per hour faster in flocks than when they were flying alone. 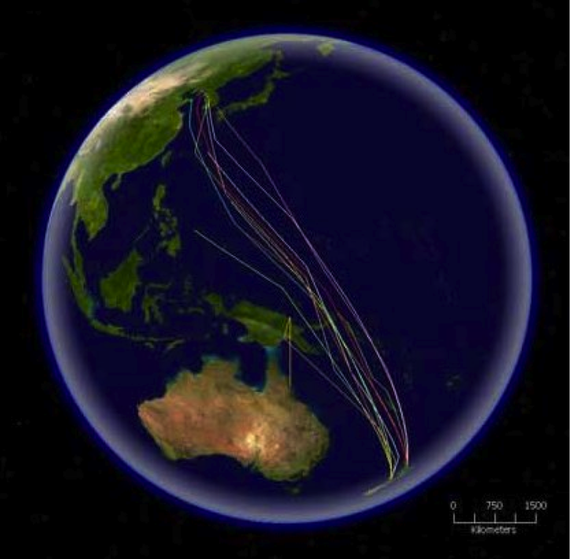 The routes of satellite tagged Bar-tailed Godwits migrating north from New Zealand, http://en.wikipedia.org/wiki/Bird_migration Birds fly at varying altitudes during migration. An expedition to Mt. Everest found skeletons of Pintail and Black-tailed Godwit at 5000 m (16,400 ft) on the Khumbu Glacier. GPS recorded that Bar-headed Geese can fly at up to 6,540 metres while crossing the Himalayas - the highest rates of climb to altitude for any bird. Seabirds fly low over water but gain altitude when crossing land. However most bird migration is in the range of 150 m (500 ft) to 600 m (2000 ft). Aviation records from the United States show most bird collisions occur below 600 m (2000 ft) and almost none above 1800 m (6000 ft). Bird migration is not limited to birds that can fly. Most species of penguin migrate by swimming over 1000 km. Blue Grouse perform altitudinal migration mostly by walking and Emus undertakes long-distance movements on foot during droughts in Australia.  Flocks of birds assembling before migration southwards http://en.wikipedia.org/wiki/Bird_migration Structures such as power lines, wind farms and offshore oil-rigs have also been known to affect migratory birds. Habitat destruction by land use changes is the biggest threat. Several international treaties have been signed to protect migratory species. http://en.wikipedia.org/wiki/Bird_migration
Toronto is mentioned as one of the world’s most deadly cities for migratory birds, because of one million to nine million birds that die every year from run-ins with it’s buildings. Casualties of Toronto’s Urban Skies Further reading: http://www.nytimes.com/slideshow/2012/10/27/world/americas/20121028-CANADA.html?ref=americas#11 http://en.wikipedia.org/wiki/The_Birds_of_America http://en.wikipedia.org/wiki/List_of_Asian_birds http://en.wikipedia.org/wiki/List_of_African_bird Source: http://en.wikipedia.org/wiki/Bird http://en.wikipedia.org/wiki/List_of_California_birds Comments are closed.
|
|
Archives
July 2020
Categories
All
|
 RSS Feed
RSS Feed
